




ICU Management & Practice, Volume 18 - Issue 3, 2018
 PRINT OPTIMISED
PRINT OPTIMISED
Xenon and brain injury.
Xenon, a chemically inert but biologically active monatomic gas, has been applied in patients for anaesthesia/sedation, and most recently in the critical care of patients with acute ongoing neurological damage.
Following preclinical evidence that xenon has ameliorative activity in several pathobiologic pathways that are involved in central nervous system injury, xenon was shown to be effective at improving both morphology and function in a series of models of hypoxic/ischaemic injury that simulate stroke (both haemorrhagic and ischaemic), neonatal asphyxia, as well as the ischaemic-reperfusion injury that occurs in the post-cardiac arrest syndrome (PCAS).
These promising findings prompted a Phase 2 RCT that revealed that a 24-hour xenon administration during targeted temperature management (TTM) resulted in significantly less brain damage than TTM alone in PCAS. A pivotal, multicentre, Phase 3 RCT is now underway to establish the efficacy (primary endpoint is survival with good functional outcome) and safety of xenon in PCAS.
Anaesthetic and sedative agents are typically administered in perioperative settings to facilitate surgical and discomfort-inducing procedures. Because of the pleiotropic effects of these powerful drugs, investigators have trialled these agents for other indications including the use of ketamine for refractory depression (McCloud et al. 2015), dexmedetomidine to facilitate detoxification from alcohol and substance abuse disorders (Wong et al. 2015), and propofol for nausea and vomiting (Griffiths et al. 2012).
Following the market authorisation for general anaesthesia in 2007, xenon’s use has extended to organ protection because of its multifaceted cytoprotective actions (Maze 2016). In this report, we focus on the neuroprotective properties of xenon.
Neuroprotective targets in acute ongoing neurologic injury
Prior to considering the possible clinical applications of xenon, we reflect on the pathophysiologic processes that characterise the clinical conditions for which xenon’s neuroprotective properties may be exploited.
Similar, but not identical, pathophysiologic mechanisms propagate ongoing damage whether the acute neurological injury is initiated by a stroke (i.e. ischaemic and haemorrhagic), cardiac arrest, or traumatic brain injury.
Stroke
Ischaemia is the cause in ~85% of adult stroke victims. Dirnagl and colleagues (2014; 1999) have offered important insights into the potentially-modifiable processes that obtain in ischaemic stroke (Figure 1). Sudden interruption of perfusion to discrete brain regions heralds a phase of excitotoxicity due to ischaemia-induced depolarisation of glutamatergic neurons causing release of the excitatory neurotransmitter, glutamate. Activation by glutamate of its cognate receptor subtypes results in a massive influx of calcium cations that produces neuronal death through necrosis. In the subacute phase of ischaemic stroke, cell death occurs through apoptotic processes. In the later phases brain damage can be produced by inflammatory processes initiated by engagement of the innate immune response (Dirnagl et al. 1999).
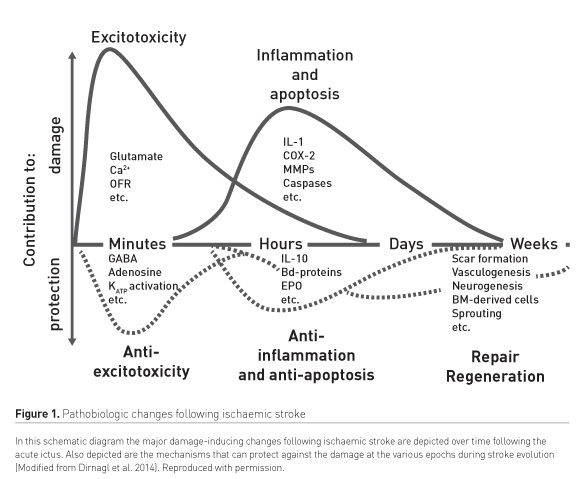
The haemorrhagic form of stroke is mainly caused by subarachnoid and intracerebral haemorrhage. The former type accounts for 5 % to 10% of all strokes and is mostly attributed to rupture of an aneurysm. Apart from the decreased perfusion and ischaemia in the territory of the ruptured blood vessel engendering excitotoxicity, additional pathophysiologic processes supervene due to the collection of extravascular blood (cytotoxic effect) (Budohoski et al. 2014). Because of perturbed cerebrospinal fluid hydrodynamics, the intracranial pressure may rise resulting in failure to adequately perfuse other brain regions. Together with vasospasm, disruption of the blood-brain barrier and the supervention of inflammation, delayed cerebral ischaemia exacerbates the patient’s neurologic deficits. However, the contribution of vasospasm to delayed cerebral injury following subarachnoid haemorrhage (SAH) has been challenged (Budohoski et al. 2014). It is notable that SAH remains an unmet treatment challenge, and novel interventions, including xenon, are worthy of consideration.
Cardiac arrest
Cardiac arrest is the classical example of ischaemic-reperfusion injury in which the absence of any perfusion to the brain provokes excitotoxicity (Neumar et al. 2008). Successful resuscitation and restoration of spontaneous circulation causes a new pathophysiologic process characterised by apoptosis and neuroinflammation.
Traumatic brain injury
Traumatic brain injury encompasses heterogeneous conditions from diverse types of trauma of varying severity; as such, different pathophysiological pathways may be involved and major international efforts are more precisely characterising the evolution of injury (International Initiative for Traumatic Brain Injury Research [InTBIR - intbir.nih.gov]; Transforming Research and Clinical Knowledge in Traumatic Brain Injury [TRACKTBI - tracktbi.ucsf.edu]). From these efforts, much has been learned about genetic factors that modulate the host-response to injury by proteins such as apolipoprotein E 4 (Lawrence et al. 2015), mitochondrial DNA haplotype (Bulstrode et al. 2014) and brain-derived neurotrophic factor (Failla et al. 2016). Physiologic monitoring has yielded information on dysregulation of intracranial pressure, autoregulation of brain perfusion, brain oxygenation and metabolism, inflammation and cortical electrical activity. Macroscopically several types of lesions can be distinguished including shearing of white matter tracts, contusions, haematomas and oedema. A secondary wave of damage occurs hours to days after the traumatic event that is characterised by excitotoxicty, free radical generation, mitochondrial dysfunction, mass effect, ischaemia and inflammatory responses (Maas et al. 2008). Especially in the setting of repetitive trauma, processes are initiated that result in long-term consequences such as dementia, Parkinsonism, and epilepsy.
Neuroprotective properties of xenon
Xenon is an antagonist of the N-methyl-D-aspartate (NMDA) subtype of the glutamate receptor (Franks et al. 1998), a pivotal mediator of the excitotoxicity that is ubiquitously present in acute ongoing neurological injury from a variety of causes. NMDA-receptor antagonists are neuroprotective in in vitro and in vivo brain injury models (Choi et al. 1988). Interventions, such as ketamine, that produce NMDA antagonism through ion pore blockade, result in the development of “Olney’s lesions” with psychotomimetic effects (Olney et al. 1991). Xenon produces its NMDA antagonism by competing with glycine at the co-activation site (Dickinson et al. 2007); hence xenon does not induce the Olney’s lesions or behavioural changes that characterise the direct ion pore blockers. In fact, xenon ameliorates the injury produced by other NMDA-receptor antagonists (Nagata et al. 2001). Xenon protects against injury induced by NMDA, glutamate or oxygen-glucose deprivation (Wilhelm et al. 2002). Other complementary neuroprotective properties of xenon include interruption of apoptosis (Ma et al. 2005), activation of species of ion channels that result in membrane hyperpolarisation (Bantel et al. 2010; Gruss et al. 2004) and a generalised cytoprotective action initiated by upregulation of hypoxia-inducible factor-1α (HIF-1α) and its downstream protective effectors (namely, erythropoietin) within the brain under normoxic conditions (Ma et al. 2009).
The neuroprotective properties of xenon have been corroborated in preclinical models of hypoxic ischemic encephalopathy (Wilhelm et al. 2002; Ma et al. 2006; Dingley 2006; Rajakumaraswamy et al. 2006; Dingley et al. 2008; Cattano et al. 2008; Valleggi et al. 2008; Luo et al. 2008; Bantel et al. 2009,) stroke (Homi et al. 2003; David et al. 2003; Limatola et al. 2010; David et al. 2010; Sheng et al. 2012), traumatic brain injury (Coburn et al. 2008; Harris et al. 2013; Campos-Pires et al. 2015,Campos-Pires et al. 2018) anaesthetic-induced developmental neurotoxicity (Ma et al. 2007; Cattano et al. 2008a; Shu et al. 2010; Cattano et al. 2011; Sabir et al. 2013), and cardiac arrest (Schmidt et al. 2005; Fries et al. 2008; Fries et al. 2012). Preclinical studies have also shown that xenon and targeted temperature management (TTM; currently, the standard of care for post-cardiac arrest syndrome) can be combined to protect in an additive or super-additive (synergistic) manner. Data, published in eight peer-reviewed manuscripts from four different laboratories involving four preclinical injury models, demonstrate that xenon’s neuroprotective action is most effective when body temperature is reduced (Ma et al. 2005; Martin et al. 2007; Hobbs et al. 2008; Thoresen et al. 2009; Chakkarapani et al. 2010; Faulkner et al. 2011; Fries et al. 2012; Sabir et al. 2014). Unlike other neuroprotective strategies, including a different NMDA-receptor antagonist (gavestinel), xenon alone exhibits this enhanced efficacy when temperature is reduced.
Clinical evidence for the neuroprotective properties of xenon
The Xe-hypotheca trial (NCT 00879892 - clinicaltrials.gov/ct2/show/NCT00879892) studied the Effect of Inhaled Xenon on Cerebral White Matter Damage in Comatose Survivors following an Out-of-Hospital Cardiac Arrest (Laitio et al. 2016). The trial, which was undertaken at the medical centres of Turku and Helsinki Universities in Finland, enrolled 110 successfully resuscitated (restoration of spontaneous circulation [ROSC] within 45 minutes of a witnessed cardiac arrest with an initial shockable rhythm) but still comatose patients. The trial compared xenon (up to 50% by inhalation) plus the standard of care (SOC) versus SOC alone for the primary endpoint of white matter brain damage (global fractional anisotropy [GFA] derived from a diffusion tensor imaging sequence); both arms were inclusive of TTM administered for 24 hrs. Assessment of the GFA revealed significantly (P=0.006) reduced brain damage in the subjects randomised to receive xenon. There was 41.7% less damage to white matter tracts (from the approximately 115,000 voxels assessed in each patient) in the Xenon + SOC vs SOC. alone group. The relative damage to the major white matter tracts in the two groups is depicted (Figure 2).
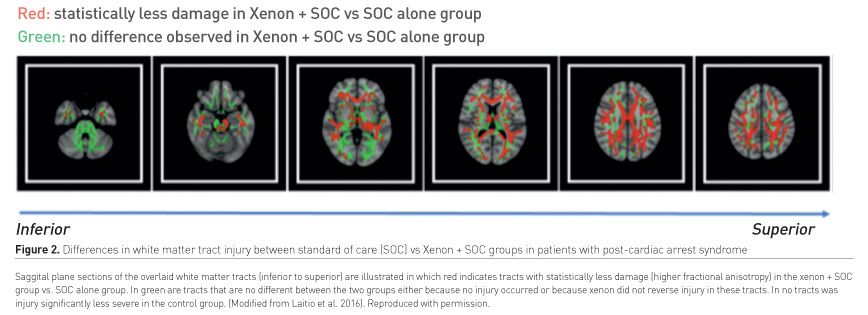
Xe-HYPOTHECA was not powered to detect differences in functional endpoints; reduction in 6-month mortality rate - 27% in the xenon group and 35% in the SOC group (adjusted hazard ratio, 0.49 [95% CI, 0.23-1.01]) - did not achieve statistical significance (P = 0.053). The degree of white matter injury was the strongest predictor of mortality at 6 months.
Clinical evidence for the myocardial protective properties of xenon
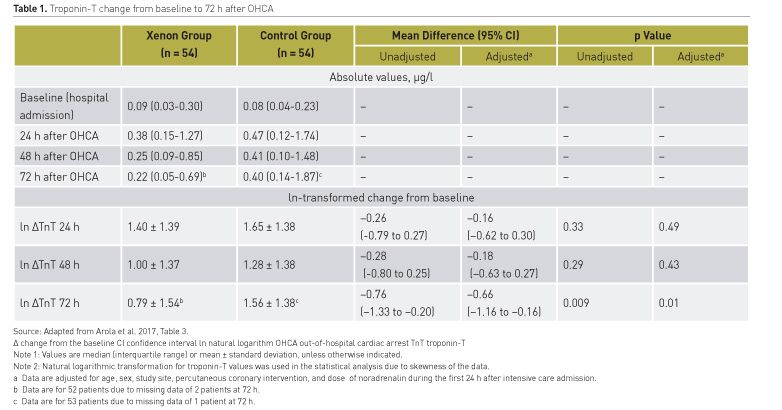
A predefined secondary objective of the XeHypotheCA Trial was to assess the effect of inhaled xenon on myocardial ischaemic damage. Troponin-T (TnT) levels were measured at hospital admission, and at 24h, 48h and 72h post-cardiac arrest (Arola et al. 2017). The baseline characteristics did not differ significantly between the groups. Results are tabulated (Table 1). After adjustments for age, gender, study site, primary coronary percutaneous intervention (PCI), and norepinephrine dose, the mean standard deviation post-arrival incremental change of the ln-transformed troponin-T at 72 hours was 0.79 (1.54) in the xenon group and 1.56 (1.38) in the control group (adjusted mean difference, -0.66 [95% CI, -1.16 ― -0.16], P=0.01). The decline of TnT from the peak value at 24h to 72h was significantly greater in the xenon group than in the control group (p =0.0008). The effect of xenon on the change in the troponin-T values did not differ in patients with or without PCI or in those with a diagnosis of ST-elevation myocardial infarction (group by PCI or STEMI interaction effect, P=0.86 and P=0.71, respectively). In comparison with hypothermia alone, inhaled xenon combined with hypothermia resulted in less severe myocardial injury as demonstrated by the significantly reduced release of troponin-T.
Future applications of xenon for critical care
Apart from the potential use of xenon for postoperative sedation (Bedi et al. 2003), and for selected intraoperative settings, e.g., for neurosurgical procedures (Rylova and Maze 2018) it is unlikely that xenon will be considered for routine use in perioperative settings both because of the availability of cheaper alternatives and because of the need for recirculating systems to minimise consumption of xenon.
Nonetheless, in critical care settings where there is an unmet medical need, e.g., Post-Cardiac Arrest Syndrome, xenon’s cytoprotective effects may be further appreciated for its physico-chemical properties that result in a near instantaneous onset of action together with a low potential for toxicity through its chemical non-reactivity. A pivotal 1436-patient Phase 3 Trial (Xenon for Neuroprotection During Post-Cardiac Arrest Syndrome in Comatose Survivors of an Out of Hospital Cardiac Arrest (XePOHCAS - NCT03176186 - clinicaltrials.gov/ct2/show/NCT03176186]) has been launched to determine the efficacy and safety of 24h inhalation of xenon to improve survival with good functional outcome in successfully resuscitated (ROSC ≤ 30 min), but still comatose, victims of a witnessed cardiac arrest.
Production and availability of xenon
Xenon is an extremely rare element present in the atmosphere at approximately 88 parts/billion. Xenon is produced by a process of cryogenic distillation in which air is fractionated into its primary components by cooling at high pressure until it liquefies; the different components are then separated according to physical characteristics, including boiling points and density, within specialised cryogenic columns. The process consumes large amounts of energy and those fractions which are found in low concentrations (i.e., xenon) require multiple distillations, and are therefore extremely expensive to produce to medical grade purity of 99.999%. The estimated total supply of xenon is thought to be 14 million litres, of which 50% could theoretically be delivered in medical grade. Approximately 50L of xenon is required for a 24-hour intervention; under these conditions approximately 140,000 patients could be treated annually.
Conclusions
Xenon, the noble elemental gas, may benefit the critically ill patient that has ongoing acute neurological injury because it reduces the activity in several of the pathobiologic pathways that obtain in these conditions. A definitive, pivotal, multicentre, trial to establish xenon’s safety and efficacy in the setting of post-cardiac arrest syndrome is now being prosecuted. If successful, the next challenge will be to increase the production of medical grade xenon through the retrofitting of oxygen-purification plants. Further refinements in the recirculation and recycling of xenon can further improve the availability of this scarce resource.
Conflict of interest
Mervyn Maze is a co-founder, and shareholder of equity in NPXe Ltd, a company that seeks to commercialise neuroprotective applications of xenon. TL is a member of the Trial Executive Committee for XePOHCAS, a phase 3 RCT designed to test the efficacy and safety of patients with post-cardiac arrest syndrome. The XePOHCAS trial is sponsored by NeuroproteXeon.
References:
Arola O, Saraste A, Laitio R et al. (2017) inhaled xenon attenuates myocardial damage in comatose survivors of out-of-hospital cardiac arrest: the Xe-Hypotheca trial. J Am Coll Cardiol, 70(21): 2652-60.
Bantel C, Maze M, Trapp S (2009) Neuronal preconditioning by inhalational anesthetics: evidence for the role of plasmalemmal adenosine triphosphate-sensitive potassium channels. Anesthesiology, 110(5): 986-95.
Bantel C, Maze M, Trapp S et al. (2010) Noble gas xenon is a novel adenosine triphosphate-sensitive potassium channel opener, Anesthesiology, 112(30: 623.
Bedi A, Murray JM, Dingley J et al. (2003) Use of xenon as a sedative for patients receiving critical care, Cri Care Med, 31(10): 2470.
Budohoski KP, Guilfoyle M, Helmy A et al. (2014) The pathophysiology and treatment of delayed cerebral ischaemia following subarachnoid haemorrhage, J Neurol Neurosurg Psychiatry, 85(12): 1343.
Bulstrode H, Nicoll JA, Hudson G et al. (2014) Mitochondrial DNA and traumatic brain injury, Ann Neurol, 75(2): 186.
Campos-Pires R, Armstrong SP, Sebastiani A et al. (2015) Xenon improves neurologic outcome and reduces secondary injury following trauma in an in vivo model of traumatic brain injury. Crit Care Med, 43(1): 149-58.
Campos-Pires R, Koziakova M, Yonis A et al. (2018) Xenon protects against blast-induced traumatic brain injury in an in vitro model. J Neurotrauma, 35(8): 1037-44.
Cattano D, Valleggi S, Ma D et al. (2008) Xenon induces transcription of ADNP in neonatal rat brain. Neurosci Lett, 440(3): 217-21.
Cattano D, Williamson P, Fukui K et al. (2008a) Potential of xenon to induce or to protect against neuroapoptosis in the developing mouse brain. Can J Anaesth, 55(7): 429-36.
Cattano D, Valleggi S, Cavazzana AO et al. (2011) Xenon exposure in the neonatal rat brain: effects on genes that regulate apoptosis. Minerva Anestesiol, 77(6): 571-8.
Chakkarapani E, Dingley J, Liu X et al. (2010) Xenon enhances hypothermic neuroprotection in asphyxiated newborn pigs. Ann Neurol, 68(3): 330-41.
Choi DW, Koh JY, Peters S (1988) Pharmacology of glutamate neurotoxicity in cortical cell culture: attenuation by NMDA antagonists. J Neurosci, 8(1): 185-96.
Coburn M, Maze M, Franks NP (2008) The neuroprotective effects of xenon and helium in an in vitro model of traumatic brain injury. Crit Care Med, 36(2): 588-95.
David HN, Leveille F, Chazalviel L et al. (2003) Reduction of ischemic brain damage by nitrous oxide and xenon. J Cereb Blood Flow Metab, 23(10): 1168-73.
David HN, Haelewyn B, Risso JJ et al. (2010) Xenon is an inhibitor of tissue-plasminogen activator: adverse and beneficial effects in a rat model of thromboembolic stroke. J Cereb Blood Flow Metab, 30(4): 718-28.
Dickinson R, Peterson R, Banks P et al. (2007) Competitive inhibition at the glycine site of the N-methyl-D-aspartate receptor by the anesthetics xenon and isoflurane: evidence from molecular modeling and electrophysiology, Anesthesiology, 107(5): 756.
Dingley J, Tooley J, Porter H et al. (2006) Xenon provides short-term neuroprotection in neonatal rats when administered after hypoxia-ischemia, Stroke, 37(2): 501.
Dingley J, Hobbs C, Ferguson J et al. (2008) Xenon/hypothermia neuroprotection regimes in spontaneously breathing neonatal rats after hypoxic-ischemic insult: the respiratory and sedative effects. Anesth Analg, 106(3): 916-23.
Dirnagl U, Iadecola C, Moskowitz MA(1999) Pathobiology of ischaemic stroke: an integrated view. Trends Neurosci, 22(9): 391-7.
Dirnagl U, Endres M (2014) Found in translation: preclinical stroke research predicts human pathophysiology, clinical phenotypes, and therapeutic outcomes, Stroke, 45(5): 1510.
Failla MD, Conley YP, Wagner AK et al. (2016) Brain-Derived Neurotrophic Factor (BDNF) in Traumatic Brain Injury-Related Mortality: Interrelationships Between Genetics and Acute Systemic and Central Nervous System BDNF Profiles, Neurorehabil Neural Repair, 30(1): 83.
Faulkner S, Bainbridge A, Kato T et al. (2011) Xenon augmented hypothermia reduces early lactate/N-acetylaspartate and cell death in perinatal asphyxia. Ann Neurol, 70(1): 133-50.
Franks NP, Dickinson R, de Sousa SL et al. (1998) How does xenon produce anaesthesia?Nature, 396(6709): 324.
Fries M, Nolte KW, Coburn M et al. (2008) Xenon reduces neurohistopathological damage and improves the early neurological deficit after cardiac arrest in pigs. Crit Care Med, 36(8): 2420-6.
Fries M, Brücken A, Çizen A et al. (2012) Combining xenon and mild therapeutic hypothermia preserves neurological function after prolonged cardiac arrest in pigs. Crit Care Med, 40(4): 1297-303.
Griffiths JD, Gyte GM, Paranjothy S et al. (2012) Interventions for preventing nausea and vomiting in women undergoing regional anaesthesia for caesarean section, Cochrane Database Syst Rev, (9).
Gruss M, Bushell TJ, Bright DP et al. (2004) Two-pore-domain K+ channels are a novel target for the anesthetic gases xenon, nitrous oxide, and cyclopropane, Mol Pharmacol, 65(2): 443.
Harris K, Armstrong SP, Campos-Pires R et al. (2013) Neuroprotection against traumatic brain injury by xenon, but not argon, is mediated by inhibition at the N-methyl-D-aspartate receptor glycine site. Anesthesiology, 119(5): 1137-48.
Hobbs C, Thoresen M, Tucker A et al. (2008) Xenon and hypothermia combine additively, offering long-term functional and histopathologic neuroprotection after neonatal hypoxia/ischemia. Stroke, 39(4): 1307-13.
Homi HM, Yokoo N, Ma D et al. (2003) The neuroprotective effect of xenon administration during transient middle cerebral artery occlusion in mice. Anesthesiology, 99(4): 876-81.
Laitio R, Hynninen M, Arola O et al. (2016) Effect of Inhaled Xenon on Cerebral White Matter Damage in Comatose Survivors of Out-of-Hospital Cardiac Arrest: A Randomized Clinical Trial. JAMA, 315(11): 1120-8.
Lawrence DW, Comper P, Hutchison MG et al. (2015) The role of apolipoprotein E episilon (ε)-4 allele on outcome following traumatic brain injury: A systematic review, Brain Inj, 29(9): 1018.
Limatola V, Ward P, Cattano D et al. (2010) Xenon preconditioning confers neuroprotection regardless of gender in a mouse model of transient middle cerebral artery occlusion. Neuroscience, 165(3): 874-81.
Luo Y, Ma D, Ieong E et al. (2008) Xenon and sevoflurane protect against brain injury in a neonatal asphyxia model.
Anesthesiology, 109(5): 782-9.
Ma D, Hossain M, Chow A et al. (2005) Xenon and hypothermia combine to provide neuroprotection from neonatal asphyxia, Ann Neurol, 58(2): 182.
Ma D, Hossain M, Pettet GK et al. (2006) Xenon preconditioning reduces brain damage from neonatal asphyxia in rats, J Cereb Blood Flow Metab, 26(2): 199.
Ma D, Williamson P, Januszewski A et al. (2007) Xenon mitigates isoflurane-induced neuronal apoptosis in the developing rodent brain. Anesthesiology, 106(4): 746-53.
Ma D, Lim T, Xu J et al. (2009) Xenon preconditioning protects against renal ischemic-reperfusion injury via HIF-1alpha activation, J Am Soc Nephrol, 20(4): 7.
Maas AL, Stocchetti N, Bullock R et al. (2008) Moderate and severe traumatic brain injury in adults, Lancet Neurol, 7(8): 728.
Martin JL, Ma D, Hossain M et al. (2007) Asynchronous administration of xenon and hypothermia significantly reduces brain infarction in the neonatal rat. Br J Anaesth, 98(2): 236-40.
Maze M (2016) Preclinical neuroprotective actions of xenon and possible implications for human therapeutics: a narrative review, Can J Anaesth, 63(2): 212.
McCloud TL, Caddy C, Jochim J et al. (2015) Ketamine and other glutamate receptor modulators for depression in bipolar disorder in adults, Cochrane Database Syst Rev, (9).
Nagata A, Nakao Si S, Nishizawa N et al. (2001) Xenon inhibits but N(2)O enhances ketamine-induced c-Fos expression in the rat posterior cingulate and retrosplenial cortices, Anesth Analg, 92(2): 362.
Neumar RW, Nolan JP, Adrie C et al. (2008) Post-cardiac arrest syndrome: epidemiology, pathophysiology, treatment, and prognostication. A consensus statement from the International Liaison Committee on Resuscitation (American Heart Association, Australian and New Zealand Council on Resuscitation, European Resuscitation Council, Heart and Stroke Foundation of Canada, InterAmerican Heart Foundation, Resuscitation Council of Asia, and the Resuscitation Council of Southern Africa); the American Heart Association Emergency Cardiovascular Care Committee; the Council on Cardiovascular Surgery and Anesthesia; the Council on Cardiopulmonary, Perioperative, and Critical Care; the Council on Clinical Cardiology; and the Stroke Council, Circulation, 118(23): 2452.
Olney JW, Labruyere J, Wang G et al. (1991) NMDA antagonist neurotoxicity: mechanism and prevention, Science, 254(5037): 1515.
Rajakumaraswamy N, Ma D, Hossain M et al. (2006), Neuroprotective interaction produced by xenon and dexmedetomidine on in vitro and in vivo neuronal injury models, Neurosci Lett, 409(2): 128.
Rylova A, Maze M (2018) Protecting the Brain with Xenon Anesthesia for Neurosurgical Procedures, J Neurosurg Anesthesiol.
Sabir H, Bishop S, Cohen N et al. (2013) Neither xenon nor fentanyl induces neuroapoptosis in the newborn pig brain. Anesthesiology, 119(2): 345-57.
Sabir H, Walløe L, Dingley J et al. (2014) Combined treatment of xenon and hypothermia in newborn rats--additive or synergistic effect? PLoS One, 9(10): e109845.
Schmidt M, Marx T, Glöggl E et al. (2005) Xenon attenuates cerebral damage after ischemia in pigs. Anesthesiology, 102(5): 929-36.
Sheng SP, Lei B, James ML et al. (2012) Xenon neuroprotection in experimental stroke: interactions with hypothermia and intracerebral hemorrhage. Anesthesiology, 117(6): 1262-75.
Shu Y, Patel SM, Pac-Soo C et al. (2010) Xenon pretreatment attenuates anesthetic-induced apoptosis in the developing brain in comparison with nitrous oxide and hypoxia. Anesthesiology, 113(2): 360-8.
Thoresen M, Hobbs CE, Wood T et al. (2009) Cooling combined with immediate or delayed xenon inhalation provides equivalent long-term neuroprotection after neonatal hypoxia-ischemia. J Cereb Blood Flow Metab, 29(4): 707-14.
Valleggi S, Cavazzana AO, Bernardi R et al. (2008) Xenon up-regulates several genes that are not up-regulated by nitrous oxide. J Neurosurg Anesthesiol, 20(4): 226-32.
Wilhelm S, Ma D, Maze M et al. (2002) Effects of xenon on in vitro and in vivo models of neuronal injury, Anesthesiology, 96(6): 1485.
Wong A, Smithburger PL, Kane-Gill SL et al. (2015) Review of adjunctive dexmedetomidine in the management of severe acute alcohol withdrawal syndrome, Am J Drug Alcohol Abuse, 41(5): 382.
Bantel C, Maze M, Trapp S (2009) Neuronal preconditioning by inhalational anesthetics: evidence for the role of plasmalemmal adenosine triphosphate-sensitive potassium channels. Anesthesiology, 110(5): 986-95.
Bantel C, Maze M, Trapp S et al. (2010) Noble gas xenon is a novel adenosine triphosphate-sensitive potassium channel opener, Anesthesiology, 112(30: 623.
Bedi A, Murray JM, Dingley J et al. (2003) Use of xenon as a sedative for patients receiving critical care, Cri Care Med, 31(10): 2470.
Budohoski KP, Guilfoyle M, Helmy A et al. (2014) The pathophysiology and treatment of delayed cerebral ischaemia following subarachnoid haemorrhage, J Neurol Neurosurg Psychiatry, 85(12): 1343.
Bulstrode H, Nicoll JA, Hudson G et al. (2014) Mitochondrial DNA and traumatic brain injury, Ann Neurol, 75(2): 186.
Campos-Pires R, Armstrong SP, Sebastiani A et al. (2015) Xenon improves neurologic outcome and reduces secondary injury following trauma in an in vivo model of traumatic brain injury. Crit Care Med, 43(1): 149-58.
Campos-Pires R, Koziakova M, Yonis A et al. (2018) Xenon protects against blast-induced traumatic brain injury in an in vitro model. J Neurotrauma, 35(8): 1037-44.
Cattano D, Valleggi S, Ma D et al. (2008) Xenon induces transcription of ADNP in neonatal rat brain. Neurosci Lett, 440(3): 217-21.
Cattano D, Williamson P, Fukui K et al. (2008a) Potential of xenon to induce or to protect against neuroapoptosis in the developing mouse brain. Can J Anaesth, 55(7): 429-36.
Cattano D, Valleggi S, Cavazzana AO et al. (2011) Xenon exposure in the neonatal rat brain: effects on genes that regulate apoptosis. Minerva Anestesiol, 77(6): 571-8.
Chakkarapani E, Dingley J, Liu X et al. (2010) Xenon enhances hypothermic neuroprotection in asphyxiated newborn pigs. Ann Neurol, 68(3): 330-41.
Choi DW, Koh JY, Peters S (1988) Pharmacology of glutamate neurotoxicity in cortical cell culture: attenuation by NMDA antagonists. J Neurosci, 8(1): 185-96.
Coburn M, Maze M, Franks NP (2008) The neuroprotective effects of xenon and helium in an in vitro model of traumatic brain injury. Crit Care Med, 36(2): 588-95.
David HN, Leveille F, Chazalviel L et al. (2003) Reduction of ischemic brain damage by nitrous oxide and xenon. J Cereb Blood Flow Metab, 23(10): 1168-73.
David HN, Haelewyn B, Risso JJ et al. (2010) Xenon is an inhibitor of tissue-plasminogen activator: adverse and beneficial effects in a rat model of thromboembolic stroke. J Cereb Blood Flow Metab, 30(4): 718-28.
Dickinson R, Peterson R, Banks P et al. (2007) Competitive inhibition at the glycine site of the N-methyl-D-aspartate receptor by the anesthetics xenon and isoflurane: evidence from molecular modeling and electrophysiology, Anesthesiology, 107(5): 756.
Dingley J, Tooley J, Porter H et al. (2006) Xenon provides short-term neuroprotection in neonatal rats when administered after hypoxia-ischemia, Stroke, 37(2): 501.
Dingley J, Hobbs C, Ferguson J et al. (2008) Xenon/hypothermia neuroprotection regimes in spontaneously breathing neonatal rats after hypoxic-ischemic insult: the respiratory and sedative effects. Anesth Analg, 106(3): 916-23.
Dirnagl U, Iadecola C, Moskowitz MA(1999) Pathobiology of ischaemic stroke: an integrated view. Trends Neurosci, 22(9): 391-7.
Dirnagl U, Endres M (2014) Found in translation: preclinical stroke research predicts human pathophysiology, clinical phenotypes, and therapeutic outcomes, Stroke, 45(5): 1510.
Failla MD, Conley YP, Wagner AK et al. (2016) Brain-Derived Neurotrophic Factor (BDNF) in Traumatic Brain Injury-Related Mortality: Interrelationships Between Genetics and Acute Systemic and Central Nervous System BDNF Profiles, Neurorehabil Neural Repair, 30(1): 83.
Faulkner S, Bainbridge A, Kato T et al. (2011) Xenon augmented hypothermia reduces early lactate/N-acetylaspartate and cell death in perinatal asphyxia. Ann Neurol, 70(1): 133-50.
Franks NP, Dickinson R, de Sousa SL et al. (1998) How does xenon produce anaesthesia?Nature, 396(6709): 324.
Fries M, Nolte KW, Coburn M et al. (2008) Xenon reduces neurohistopathological damage and improves the early neurological deficit after cardiac arrest in pigs. Crit Care Med, 36(8): 2420-6.
Fries M, Brücken A, Çizen A et al. (2012) Combining xenon and mild therapeutic hypothermia preserves neurological function after prolonged cardiac arrest in pigs. Crit Care Med, 40(4): 1297-303.
Griffiths JD, Gyte GM, Paranjothy S et al. (2012) Interventions for preventing nausea and vomiting in women undergoing regional anaesthesia for caesarean section, Cochrane Database Syst Rev, (9).
Gruss M, Bushell TJ, Bright DP et al. (2004) Two-pore-domain K+ channels are a novel target for the anesthetic gases xenon, nitrous oxide, and cyclopropane, Mol Pharmacol, 65(2): 443.
Harris K, Armstrong SP, Campos-Pires R et al. (2013) Neuroprotection against traumatic brain injury by xenon, but not argon, is mediated by inhibition at the N-methyl-D-aspartate receptor glycine site. Anesthesiology, 119(5): 1137-48.
Hobbs C, Thoresen M, Tucker A et al. (2008) Xenon and hypothermia combine additively, offering long-term functional and histopathologic neuroprotection after neonatal hypoxia/ischemia. Stroke, 39(4): 1307-13.
Homi HM, Yokoo N, Ma D et al. (2003) The neuroprotective effect of xenon administration during transient middle cerebral artery occlusion in mice. Anesthesiology, 99(4): 876-81.
Laitio R, Hynninen M, Arola O et al. (2016) Effect of Inhaled Xenon on Cerebral White Matter Damage in Comatose Survivors of Out-of-Hospital Cardiac Arrest: A Randomized Clinical Trial. JAMA, 315(11): 1120-8.
Lawrence DW, Comper P, Hutchison MG et al. (2015) The role of apolipoprotein E episilon (ε)-4 allele on outcome following traumatic brain injury: A systematic review, Brain Inj, 29(9): 1018.
Limatola V, Ward P, Cattano D et al. (2010) Xenon preconditioning confers neuroprotection regardless of gender in a mouse model of transient middle cerebral artery occlusion. Neuroscience, 165(3): 874-81.
Luo Y, Ma D, Ieong E et al. (2008) Xenon and sevoflurane protect against brain injury in a neonatal asphyxia model.
Anesthesiology, 109(5): 782-9.
Ma D, Hossain M, Chow A et al. (2005) Xenon and hypothermia combine to provide neuroprotection from neonatal asphyxia, Ann Neurol, 58(2): 182.
Ma D, Hossain M, Pettet GK et al. (2006) Xenon preconditioning reduces brain damage from neonatal asphyxia in rats, J Cereb Blood Flow Metab, 26(2): 199.
Ma D, Williamson P, Januszewski A et al. (2007) Xenon mitigates isoflurane-induced neuronal apoptosis in the developing rodent brain. Anesthesiology, 106(4): 746-53.
Ma D, Lim T, Xu J et al. (2009) Xenon preconditioning protects against renal ischemic-reperfusion injury via HIF-1alpha activation, J Am Soc Nephrol, 20(4): 7.
Maas AL, Stocchetti N, Bullock R et al. (2008) Moderate and severe traumatic brain injury in adults, Lancet Neurol, 7(8): 728.
Martin JL, Ma D, Hossain M et al. (2007) Asynchronous administration of xenon and hypothermia significantly reduces brain infarction in the neonatal rat. Br J Anaesth, 98(2): 236-40.
Maze M (2016) Preclinical neuroprotective actions of xenon and possible implications for human therapeutics: a narrative review, Can J Anaesth, 63(2): 212.
McCloud TL, Caddy C, Jochim J et al. (2015) Ketamine and other glutamate receptor modulators for depression in bipolar disorder in adults, Cochrane Database Syst Rev, (9).
Nagata A, Nakao Si S, Nishizawa N et al. (2001) Xenon inhibits but N(2)O enhances ketamine-induced c-Fos expression in the rat posterior cingulate and retrosplenial cortices, Anesth Analg, 92(2): 362.
Neumar RW, Nolan JP, Adrie C et al. (2008) Post-cardiac arrest syndrome: epidemiology, pathophysiology, treatment, and prognostication. A consensus statement from the International Liaison Committee on Resuscitation (American Heart Association, Australian and New Zealand Council on Resuscitation, European Resuscitation Council, Heart and Stroke Foundation of Canada, InterAmerican Heart Foundation, Resuscitation Council of Asia, and the Resuscitation Council of Southern Africa); the American Heart Association Emergency Cardiovascular Care Committee; the Council on Cardiovascular Surgery and Anesthesia; the Council on Cardiopulmonary, Perioperative, and Critical Care; the Council on Clinical Cardiology; and the Stroke Council, Circulation, 118(23): 2452.
Olney JW, Labruyere J, Wang G et al. (1991) NMDA antagonist neurotoxicity: mechanism and prevention, Science, 254(5037): 1515.
Rajakumaraswamy N, Ma D, Hossain M et al. (2006), Neuroprotective interaction produced by xenon and dexmedetomidine on in vitro and in vivo neuronal injury models, Neurosci Lett, 409(2): 128.
Rylova A, Maze M (2018) Protecting the Brain with Xenon Anesthesia for Neurosurgical Procedures, J Neurosurg Anesthesiol.
Sabir H, Bishop S, Cohen N et al. (2013) Neither xenon nor fentanyl induces neuroapoptosis in the newborn pig brain. Anesthesiology, 119(2): 345-57.
Sabir H, Walløe L, Dingley J et al. (2014) Combined treatment of xenon and hypothermia in newborn rats--additive or synergistic effect? PLoS One, 9(10): e109845.
Schmidt M, Marx T, Glöggl E et al. (2005) Xenon attenuates cerebral damage after ischemia in pigs. Anesthesiology, 102(5): 929-36.
Sheng SP, Lei B, James ML et al. (2012) Xenon neuroprotection in experimental stroke: interactions with hypothermia and intracerebral hemorrhage. Anesthesiology, 117(6): 1262-75.
Shu Y, Patel SM, Pac-Soo C et al. (2010) Xenon pretreatment attenuates anesthetic-induced apoptosis in the developing brain in comparison with nitrous oxide and hypoxia. Anesthesiology, 113(2): 360-8.
Thoresen M, Hobbs CE, Wood T et al. (2009) Cooling combined with immediate or delayed xenon inhalation provides equivalent long-term neuroprotection after neonatal hypoxia-ischemia. J Cereb Blood Flow Metab, 29(4): 707-14.
Valleggi S, Cavazzana AO, Bernardi R et al. (2008) Xenon up-regulates several genes that are not up-regulated by nitrous oxide. J Neurosurg Anesthesiol, 20(4): 226-32.
Wilhelm S, Ma D, Maze M et al. (2002) Effects of xenon on in vitro and in vivo models of neuronal injury, Anesthesiology, 96(6): 1485.
Wong A, Smithburger PL, Kane-Gill SL et al. (2015) Review of adjunctive dexmedetomidine in the management of severe acute alcohol withdrawal syndrome, Am J Drug Alcohol Abuse, 41(5): 382.

